AXE 1:
Étude du mécanisme de terminaison de la transcription des gènes non codants et codant pour les protéines.
(PIs: D. Libri & O. Porrua)
La transcription n'est pas limitée aux régions codant pour des ARN fonctionnels connus, mais a lieu virtuellement partout dans le génome. Ce phénomène est appelé transcription pervasive ou cachée (voir nos revues Villa & Porrua, 2022 ; et Jensen et al, 2013) et est conservé dans tous les organismes de la bactérie à l'homme.
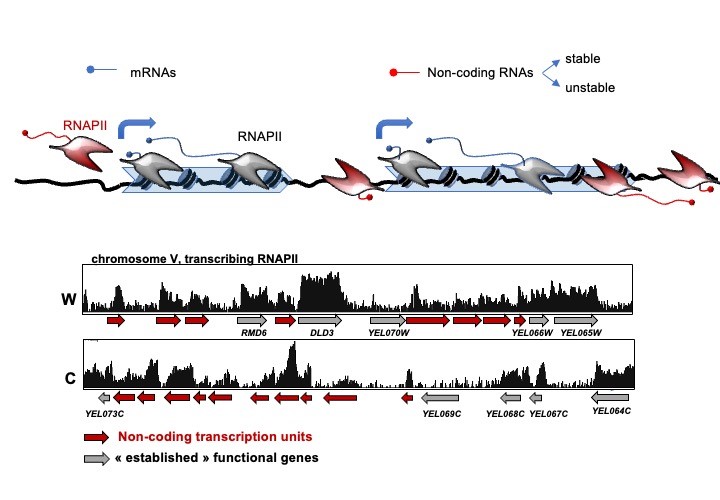
L’étendue des événements de transcription non-codants doit être contrôlée pour éviter des interférences avec l'expression des gènes voisins et d'autres processus associés à l'ADN. La terminaison de la transcription joue un rôle important dans ce contexte (voir notre revue : Porrua & Libri, 2015), et l'étude des mécanismes impliqués dans ce processus est l'un de nos principaux intérêts. La terminaison de la transcription à l'extrémité 3' des gènes codant pour des protéines dépend d'un complexe multi-sous-unités conservé appelé le complexe CPF-CF. Chez la levure bourgeonnante, le complexe NNS, composé des protéines de liaison à l'ARN Nrd1 et Nab3, et de l'hélicase Sen1, termine les événements de transcription pervasive mais aussi la transcription de gènes d'ARN non-codants fonctionnels tels que les snoRNAs. De plus, le complexe NNS favorise la dégradation des ARN non-codants par l'exosome nucléaire et son cofacteur le complexe TRAMP (figure 2).
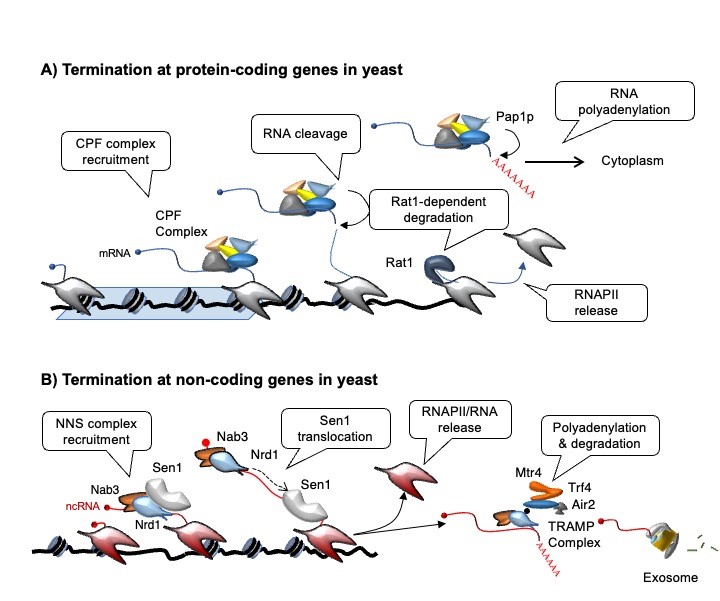
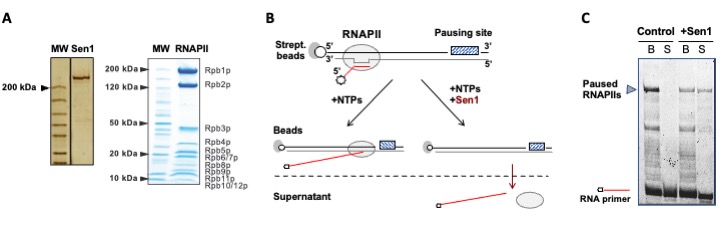
Au cours des dernières années, nous avons largement caractérisé la fonction du complexe NNS. Cependant, de nombreux aspects des mécanismes de terminaison restent obscurs, aussi bien pour les gènes non-codants que pour les gènes codant pour ARNm. Nous utilisons une technique qui permet de détecter in vivo la position de l’ARNpol II avec une résolution d'un seul nucléotide (CRAC, Crosslinking Analysis of cDNAs, figure 3, Bohnsack et al., 2012 ; Challal et al., 2022) pour générer des cartes de transcription dans des mutants du complexe NNS et de la voie CPF-CF.
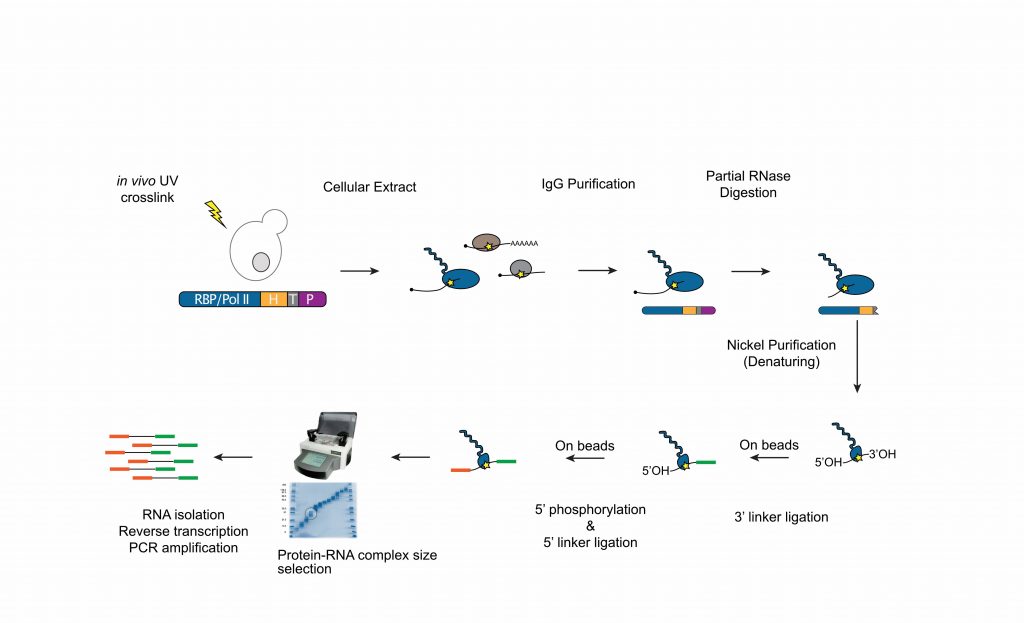
Cela permet de réaliser des analyses comparatives des différentes voies de terminaison et fournit des informations sur les mécanismes (ainsi que des résultats inattendus !). Nous aimons combiner ces analyses à haute résolution à l'échelle du génome avec des approches biochimiques in vitro (figure 4) et structurelles pour étudier les mécanismes de terminaison de la transcription et l'interaction entre les différentes voies de terminaison (comme exemple typique, voir notre publication récente : Xie et al, 2022).
AXE 2:
Analyse de l'impact de la transcription non-codante dans l'expression des gènes dans différentes conditions physiologiques
(PI : D. Libri & O. Porrua)
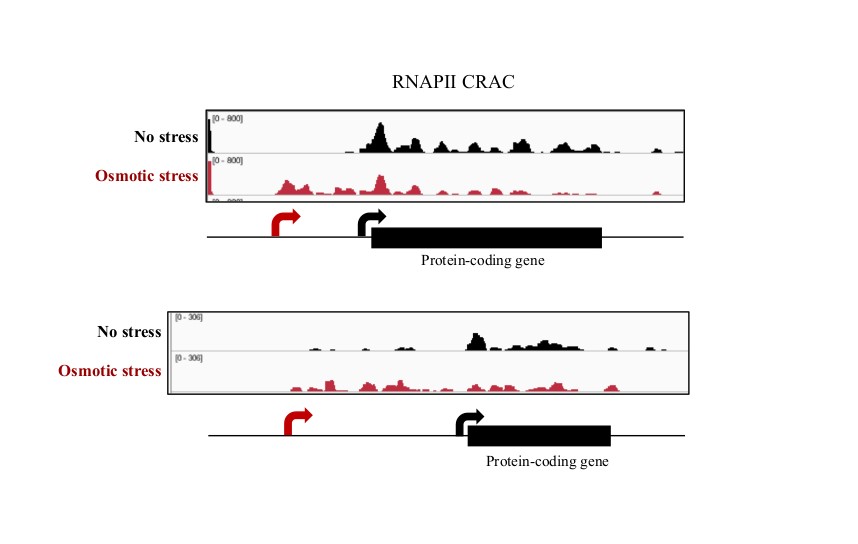
Les événements de transcription non-codante peuvent réguler l'expression des gènes en affectant la fonction des promoteurs des gènes voisins. Le potentiel régulateur de la transcription non-codante a été généralement négligé, principalement parce que de nombreuses analyses antérieures reposaient sur la détection de l'ARN comme proxy de la transcription et que les ARNnc produits par des événements de transcription potentiellement régulateurs sont souvent instables et difficilement détectables dans les cellules sauvages. La cartographie à haute résolution et directionnelle de l’ARNpol II en cours de transcription permet de contourner ce problème.
Nous sommes intéressés à explorer davantage les voies de régulation par la transcription non-codante dans différentes conditions physiologiques et de stress. Nous avons détecté de nombreux nouveaux événements de transcription d'ARNnc dans ces conditions, dont certains dérivent de promoteurs bidirectionnels activés, d'autres d'unités de transcription non-codantes "solitaires" et d'autres encore d'une efficacité réduite de la terminaison de la transcription (voir notre récente publication Haidara et al, 2022 ; et figure 5). Nous poursuivons l'étude de l'impact de la transcription non-codante sur l'expression des gènes en utilisant une variété d'approches et d'outils bioinformatiques.
AXE 3 :
Caractérisation des mécanismes responsables de la résolution des conflits transcription-réplication
(PI : D. Libri)
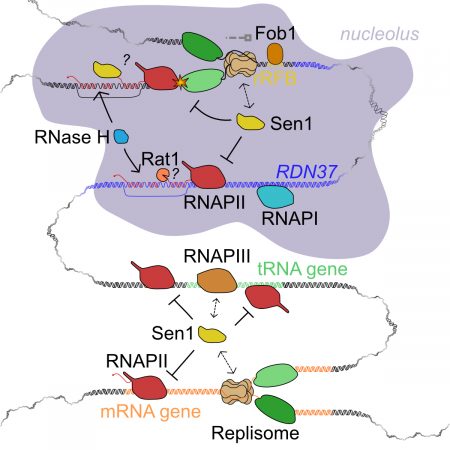
L'existence d'événements de transcription qui transcendent les limites des gènes canoniques annotés est un défi majeur pour la cohabitation de la transcription et d'autres événements associés à l'ADN comme la réplication. Nous nous sommes intéressés dans un passé récent à l'impact que la transcription pervasive a sur la fonction des origines de réplication de la levure (voir Candelli et al, 2018). Nous orientons maintenant nos intérêts vers les conflits générés par la transcription par l’ARNpol II. Dans une série d'études récentes (Aiello et al., 2022 ; Appanah et al., 2020), nous avons analysé les relations entre la transcription par l’ARNpol II et les événements de réplication et de transcription médiés par d'autres polymérases. Nous avons démontré, en collaboration avec les laboratoires de Piccoli, Pasero et Palancade, que Sen1, indépendamment de son rôle dans la terminaison de la transcription non-codante, a aussi un rôle capital dans le contrôle des conflits transcription-réplication et transcription-transcription. Nous avons montré que Sen1 élimine les ARNpols II qui entrent en collision avec le réplisome ou avec d'autres ARN polymérases en cours de transcription, se qualifiant ainsi comme un régulateur majeur de l'encombrement génomique (Figure 6).
Lorsque des conflits se produisent, l'ARN naissant peut s'hybrider au brin d'ADN matrice, formant des structures appelées R-loops. Ces structures peuvent générer des dommages à l'ADN et sont généralement éliminées par les RNases H qui dégradent la partie ARN de l'hybride. On pense également que Sen1 joue un rôle dans la limitation de la formation des R-loops ou dans leur résolution une fois qu'ils sont formés. Nous avons étudié le rôle des R-loops et des RNases H dans les régions de conflits, et décrit une nouvelle méthode de détection des R-loops à haute résolution. Nous poursuivrons les études sur le mécanisme de résolution des conflits, les rôles des RNases H et des R-loops dans ces processus et l'impact sur la stabilité du génome.
AXIS 4 :
Étude de la fonction moléculaire de la sénataxine humaine et de son implication dans les maladies neurodégénératives
(PI : O.Porrua)
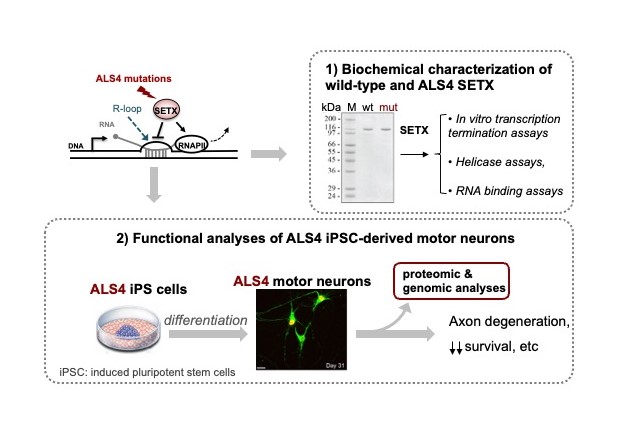
L'homologue humain de Sen1, la sénataxine (SETX), a attiré beaucoup d'attention en raison de son lien avec deux maladies neurodégénératives. Des mutations récessives de perte de fonction de SETX ont été associées à l'ataxie avec apraxie oculomotrice de type 2 (AOA2), tandis que des mutations dominantes de gain de fonction de SETX sont liées à une forme juvénile de sclérose latérale amyotrophique (SLA) appelée SLA4 (ALS4 en anglais). Comme Sen1, SETX a été attribuée un rôle dans la terminaison de la transcription ainsi que dans la résolution des R-loops, cependant le rôle précis de SETX dans ces processus restait mal compris à cause de l'absence de données biochimiques sur les propriétés et les activités de SETX. De plus, une identification systématique des protéines et ARNs interagissant avec SETX n’a pas été réalisée. Pour combler ces lacunes, nous avons mis à profit notre expertise et les outils que nous avons développés pour la caractérisation fonctionnelle de l'homologue de SETX chez la levure pour élucider la fonction moléculaire de SETX. En collaboration avec M. Sebesta et R. Stefl, nous avons récemment purifié le domaine catalytique de SETX et, en utilisant une variété d’essais biochimiques in vitro, nous avons montré pour la première fois que SETX est une hélicase capable de résoudre des R-loops et un facteur de terminaison de la transcription (voir notre pre-print Hasanova et al, 2022, https://www.biorxiv.org/content/10.1101/2022.08.25.505353v1).
De plus, nous étudions actuellement les bases moléculaires de la SLA associée à SETX en collaboration avec S. Nedelec (IFM, Paris). A cette fin, nous avons généré des motoneurones humains à partir de cellules souches pluripotentes induites portant des mutations SLA4 et nous utilisons une variété d'approches pour étudier l'impact de ces mutations sur la physiologie des motoneurones. Ensuite, nous combinerons des approches biochimiques, protéomiques et génomiques pour dévoiler les dérégulations responsables de la dégénérescence des motoneurones dans la SLA4 (figure 7).
AXIS 5 :
Dynamique de réplication de l'ADN à l’échelle de la molécule unique
(PI : M. Radman-Livaja)
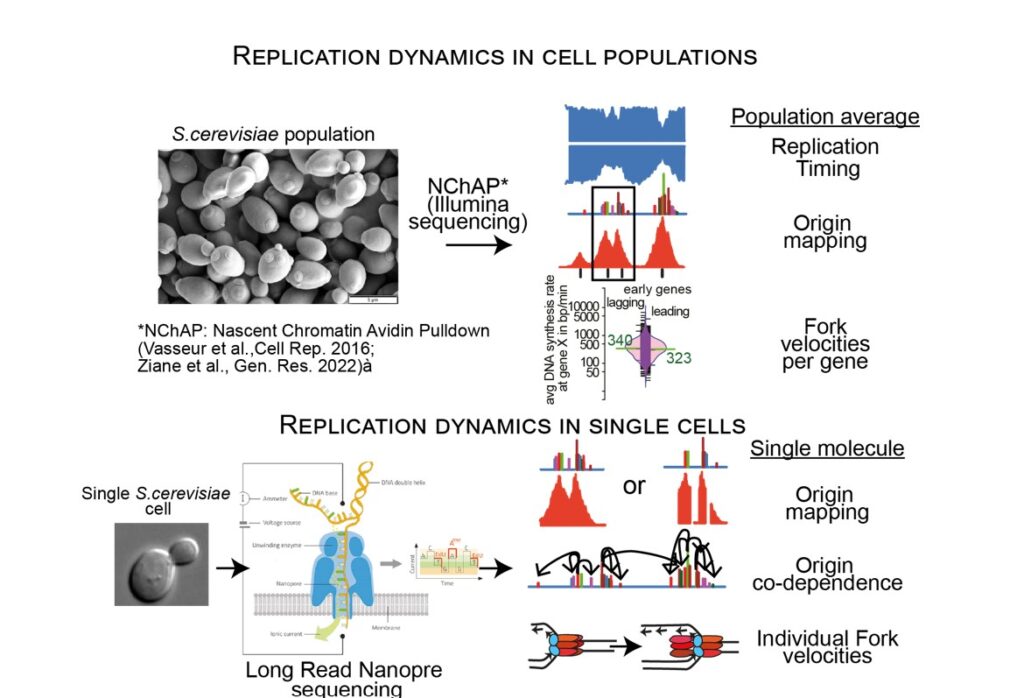
Marta Radman-Livaja a développé le NChAP (Nascent Chromatin Avidin Pulldown), une méthode de cartographie génomique spécifique à chaque brin pour étudier la dynamique de réplication de l'ADN, basée sur le séquençage Illumina de nouvelle génération (Vasseur et al., Cell Reports, 2016 ; Ziane et al., Genome Research, 2022). Le NChAP permet de produire des cartes génomiques à haute résolution du « timing » de réplication, de la localisation des origines de réplication et des vitesses de synthèse de l'ADN pour chaque gène. Ces cartes ont révélé une mine d’informations inédites, comme les différences dans le « timing » de réplication entre les brins précoces et les brins tardifs allant de 0.5 à 6min dépendant du site génomique, ou bien la présence de multiples origines densément regroupés à la place d’origines uniques précédemment identifiées.
Bien que ces cartes offrent un nouveau regard sur les mécanismes qui régissent la réplication du génome, elles ne représentent qu’une moyenne à l’échelle de la population cellulaire. Pour répondre à des questions telles que : 1) Combien d’origines sont activées dans une seule cellule? ou 2) L’activation d’une origine dépend-elle de l’activation d’autres origines sur le même chromosome ? ou encore 3) Quelle est la vitesse des fourches de réplication individuelles ?, il est nécessaire d’utiliser des technologies de séquençage de longues molécules uniques. Nous développons donc des méthodes basées sur le séquençage Nanopore pour étudier la dynamique de l’activation des origines de réplication et de la progression des fourches (y compris le suivi des conflits entre réplication et transcription) sur des brins d’ADN uniques directement issus du génome nucléaire et du génome mitochondrial de la levure bourgeonnante.