AXIS 1:
Study of the mechanism of transcription termination for non-coding and protein-coding genes
(PIs: D. Libri & O. Porrua)
Transcription is not limited to regions annotated for producing known functional RNAs, but occurs virtually everywhere in the genome, a phenomenon conserved from bacteria to humans that is named pervasive or hidden transcription (see our reviews Villa & Porrua, 2022; and Jensen et al, 2013).
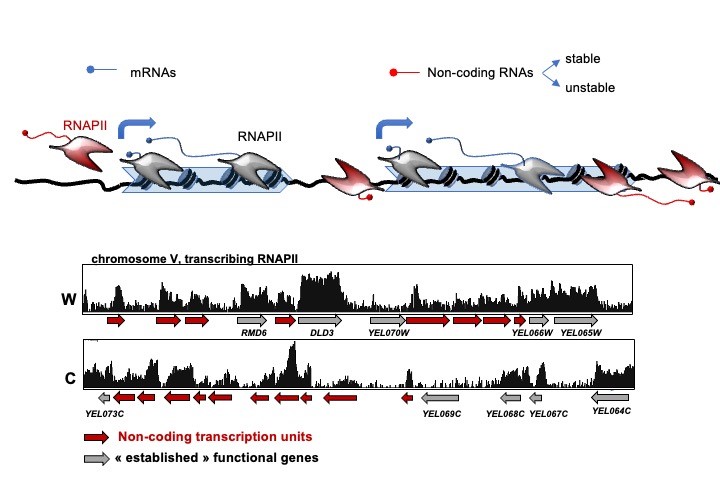
The extended distribution of non-coding transcription events needs to be controlled to avoid potentially disruptive interference with the expression of neighboring genes and other DNA-associated processes. Transcription termination has important roles in this context (see our review: Porrua & Libri, 2015), and studying the mechanisms involved in this process is one of our main interests. Transcription termination at the 3' end of protein-coding genes depends on a conserved multi-subunit complex called the CPF-CF complex. In budding yeast, the NNS complex, composed of the RNA-binding proteins Nrd1 and Nab3, and the helicase Sen1 terminates pervasive transcription events but also transcription of functional non-coding RNA genes such as snoRNAs. Moreover, the NNS complex promotes the degradation of non-coding RNAs by the nuclear exosome and its cofactor the TRAMP complex (figure 2).
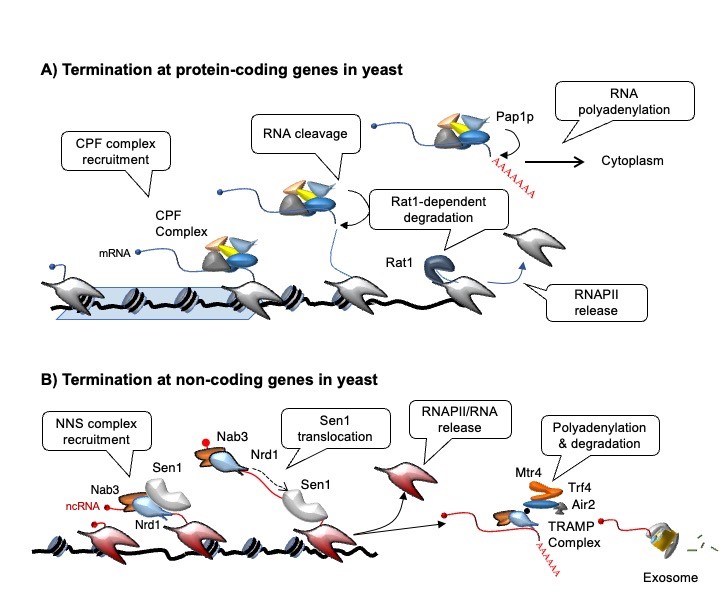
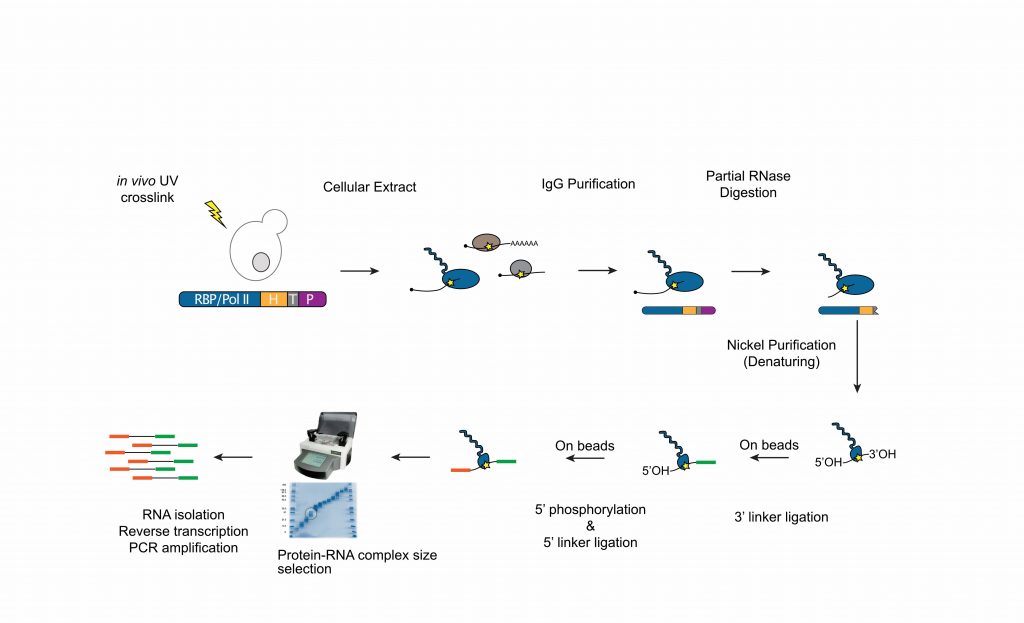
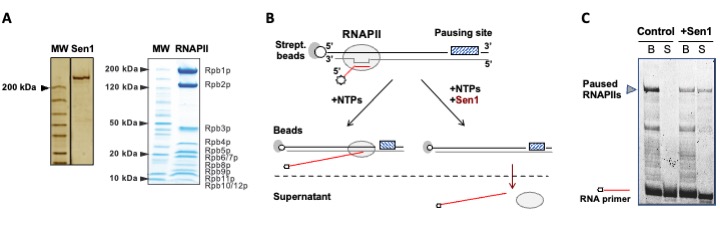
During the past years, we have extensively characterized the function of the NNS-complex. However, many aspects of the mechanisms of termination remain unclear, both for non-coding and mRNA coding genes. We use a technique that allows detecting in vivo the position of the RNAPII with single-nucleotide resolution (CRAC, Crosslinking Analysis of cDNAs, figure 3, Bohnsack et al., 2012; Challal et al., 2022) to generate transcription maps in mutants of the NNS and the CPF-CF pathways.
.
This allows comparative analyses of the different termination pathways and provides some information on the mechanism (together with some unexpected findings!). We like to combine these high-resolution genome-wide analyses with in vitro biochemical (figure 4) and structural approaches to investigate the mechanisms of transcription termination and the interplay between the different termination pathways (as a typical example see our recent publication: Xie et al, 2022).
AXIS 2:
Analysis of the impact of non-coding transcription in gene expression in different physiological conditions
(PI : D. Libri & O. Porrua)
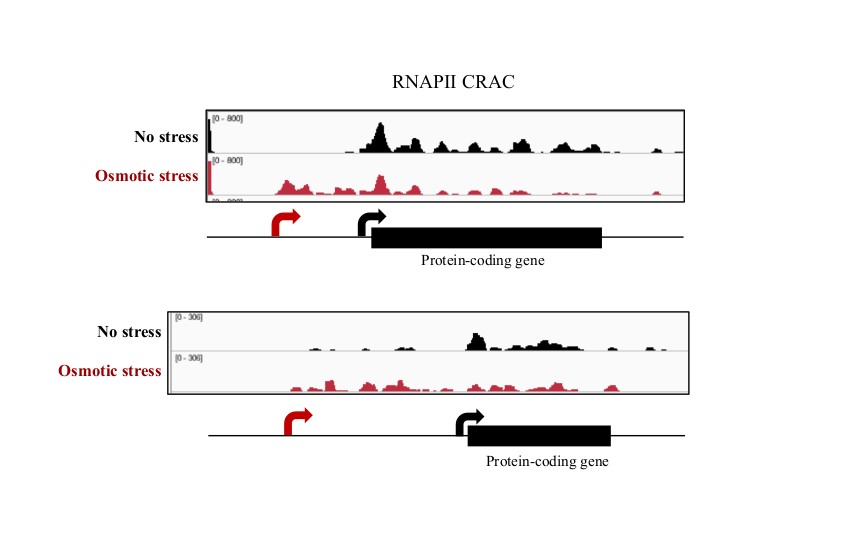
Non-coding transcription events can regulate gene expression by affecting the function of promoters of neighboring genes in the sense or antisense orientation. The regulatory potential of non-coding transcription has been generally neglected, mostly because many previous analyses relied on detection of the RNA as proxy for transcription and ncRNAs produced by potentially regulatory transcription events are often unstable and not easily detected in wild type cells. High resolution and directional mapping the transcribing RNAPII allows circumventing this problem.
We are interested in further exploring pathways of regulation by non-coding transcription in different physiological and stress conditions. We detected many novel ncRNA transcription events in these conditions, some of which derive from activated bidirectional promoters, some from “solo” non-coding transcription units and some from decreased transcription termination efficiency (see our recent publication Haidara et al, 2022; and figure 5). We are pursuing the study of the impact of non-coding transcription in gene expression using a variety of approaches and bioinformatic tools.
AXIS 3 :
Characterization of the mechanisms responsible for the resolution of transcription-replication conflicts
(PI : D. Libri)
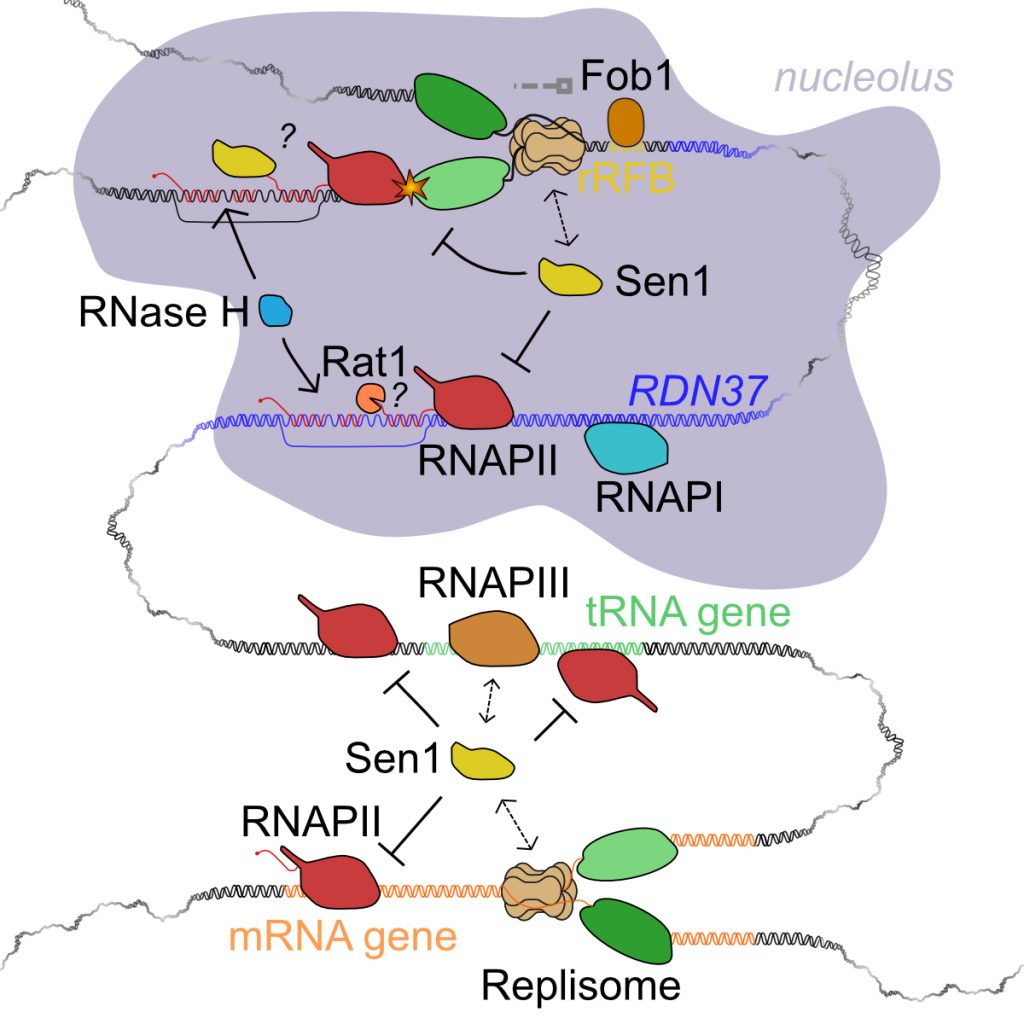
The existence of transcription events that transcend the limits of annotated canonical genes is a major challenge for the cohabitation of transcription and other DNA-associated events such as replication. We have been interested in the recent past the impact that pervasive transcription has on the function of yeast replication origins (see Candelli et al, 2018). We are now directing our interests to the conflicts that are generated by RNAPII transcription. In a series of recent studies (Aiello et al., 2022; Appanah et al., 2020) we have analyzed the relationships of RNAPII transcription with replication and transcription events mediated by other polymerases. We have demonstrated, in collaboration with the de Piccoli, Pasero and Palancade labs, that Sen1, besides and independently of its role in terminating non-coding transcription, has a capital role in controlling transcription-replication and transcription-transcription conflicts. We have shown that Sen1 removes RNAPIIs that collide with the replisome or with transcription of other polymerases, thus qualifying as a master regulator of genomic crowding (Figure 6).
When conflicts occur, the nascent RNA might hybridize to the template DNA strand, forming structures called R-loops. These structures can generate DNA damage and be genotoxic, and are generally removed by RNases H that degrade the RNA portion of the hybrid. Sen1 is also believed to play a role in limiting the formation of R-loops or resolving them once they form. We studied the role of R-loops and RNases H in regions of conflicts, and described a novel method for high resolution R-loop detection. We will pursue the studies of the mechanism of conflict resolution, the roles of RNases H and R-loops in these processes and the impact in genome stability.
AXIS 4 :
Study of the molecular function of human senataxin and its involvement in neurodegeneration
(PI : O.Porrua)
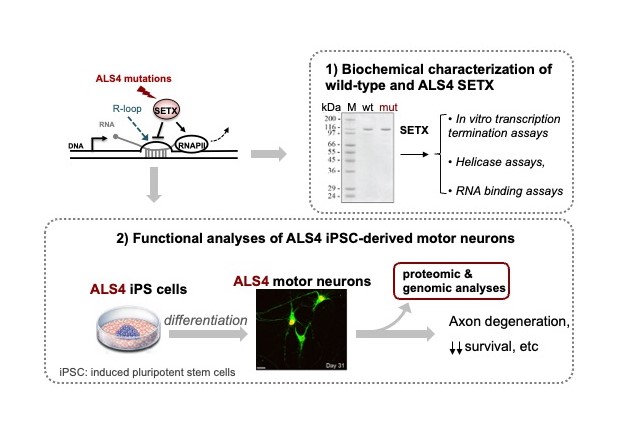
The human homologue of Sen1, senataxin (SETX), has attracted much attention because of its connection with two neurodegenerative diseases. Recessive loss-of-function SETX mutations have been associated with Ataxia with Oculomotor Apraxia type 2 (AOA2), whereas dominant gain-of-function mutations in SETX are linked to a juvenile form of Amyotrophic Lateral Sclerosis (ALS) dubbed ALS4. As Sen1, SETX has been assigned a role in transcription termination as well as in R-loop resolution, however the precise role of SETX in these processes has remained poorly understood because of the absence of biochemical data on the properties and activities of SETX. In addition, a systematic identification of the protein interactors and the targets of SETX is missing. To fill these gaps, we have benefitted from our expertise and the tools we have developed for the functional characterization of the yeast homologue of SETX to elucidate the molecular function of SETX. In collaboration with M. Sebesta and R. Stefl we have recently purified the catalytic domain of SETX and, using a variety of in vitro biochemical assays we have shown for the first time that SETX is a bona fide R-loop resolving helicase and transcription termination factors (see our recent preprint Hasanova et al, 2022, https://www.biorxiv.org/content/10.1101/2022.08.25.505353v1).
In addition, we are currently addressing the molecular basis of SETX-associated ALS in collaboration with S. Nedelec (IFM, Paris). To this end, we have generated human motor neurons from induced pluripotent stem cells harbouring ALS4 mutations and we are employing a variety of approaches to investigate the impact of these mutations on motor neuron physiology. Next, we will combine biochemical, proteomic and genomic approaches to unveil the deregulations responsible for motor neuron impairment in ALS4 (figure 7).
AXIS 5 :
Single-molecule DNA Replication Dynamics
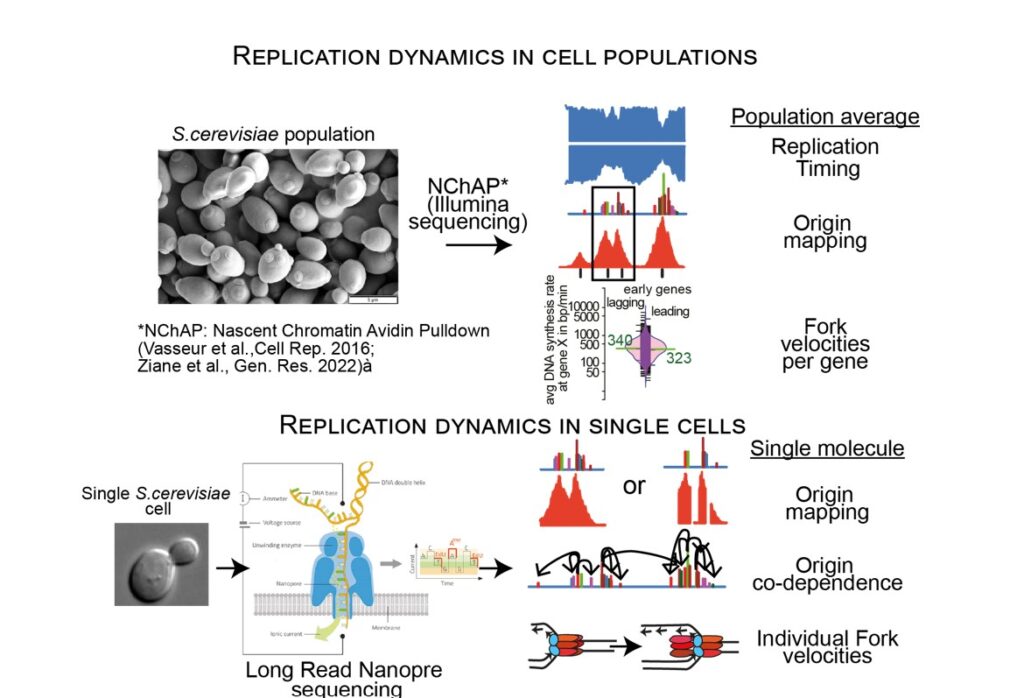
(PI : M. Radman-Livaja)
Marta Radman-Livaja has developed NChAP: (Nascent Chromatin Avidin Pulldown), a method for strand-specific genome-wide mapping of DNA replication dynamics based on Illumina NG Sequencing (Vasseur et al.,Cell. Rep., 2016; Ziane et al., Gen. Res. 2022). NChAP produces high-resolution genome-wide maps of yeast replication timing, yeast replication origins and DNA synthesis/replication fork rates. These maps revealed a trove of new information such as locus- specific differences in replication timing between the lagging and the leading strands that range from 0.5 to 6min or the presence of densely packed multi-origin clusters in place of previously identified single origins.
While these maps shed unprecedented new light on the mechanisms governing genome replication they only represent cell population averages. In order to address questions like: 1) How many origins are activated in an individual cell?; 2) Does the activation of an origin depend on the activation of other origins on the same chromosome?; or 3) What are the velocities of individual forks?, we need to use single-molecule approached. To this end, we are currently developing methods based on Nanopore long-read sequencing technology to investigate single-molecule dynamics of replication origin activation and fork progression (including monitoring of replication/transcription conflicts) on nuclear and mitochondrial genomes of budding yeast.