Selection of publications :
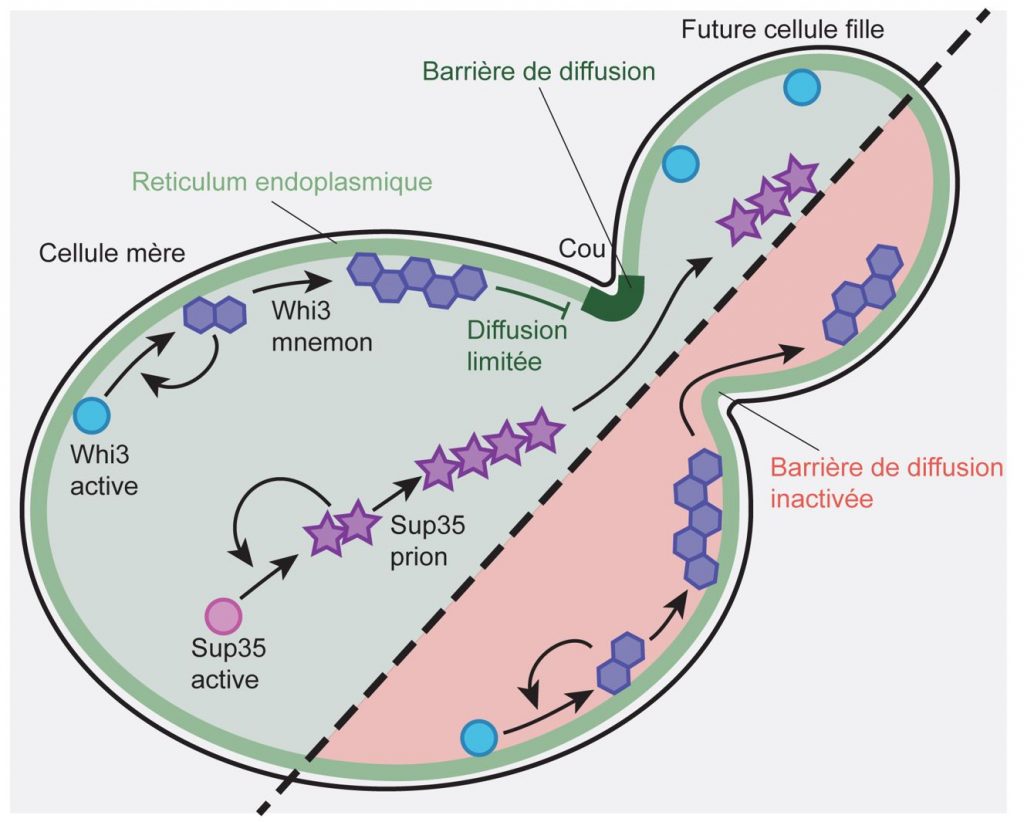
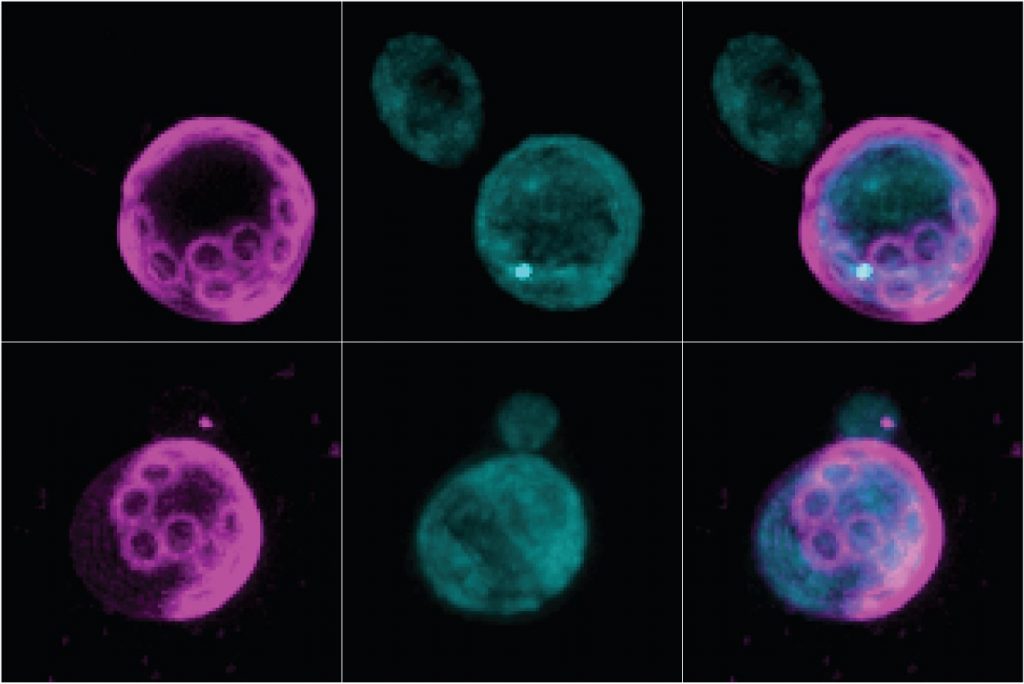
Whi3 mnemon association with endoplasmic reticulum membranes confines the memory of deceptive courtship to the yeast mother cell. Lau, Y., Oamen, H.P., Grogg, M., Parfenova, I., Saarikangas, J., Hannay, R., Nichols, R.A., Hilvert, D., Barral, Y., and Caudron, F. (2022). Current Biology.
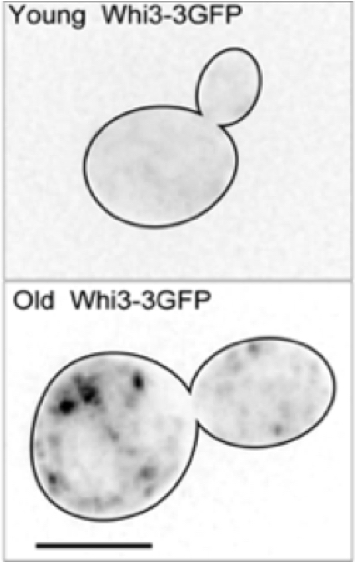
Aggregation of the Whi3 protein, not loss of heterochromatin, causes sterility in old yeast cells. Schlissel, G., Krzyzanowski, M.K., Caudron, F., Barral, Y., and Rine, J. (2017). Science 355, 1184–1187.
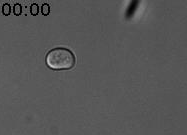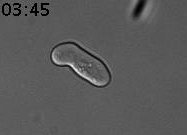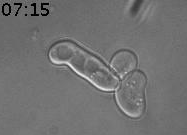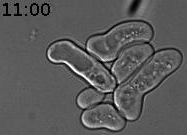
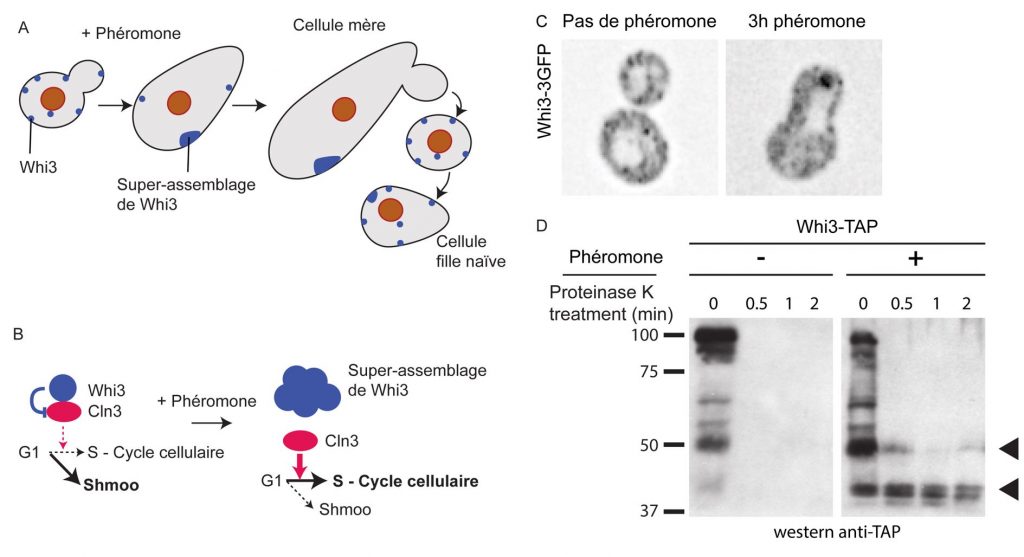
A super-assembly of Whi3 encodes memory of deceptive encounters by single cells during yeast courtship. Caudron, F., and Barral, Y. (2013). Cell 155, 1244–1257.
List of all the publications :
1. A rare natural lipid induces neuroglobin expression to prevent amyloid oligomers toxicity and retinal neurodegeneration. Oamen, H.P., Romero, N.R., Knuckles, P., Saarikangas, J., Dong, Y., and Caudron, F. (2021). bioRxiv, 20. https://doi.org/10.1101/2021.06.23.449608
2. Whi3 mnemon association with endoplasmic reticulum membranes confines the memory of deceptive courtship to the yeast mother cell. Lau, Y., Oamen, H.P., Grogg, M., Parfenova, I., Saarikangas, J., Hannay, R., Nichols, R.A., Hilvert, D., Barral, Y., and Caudron, F. (2022). Current Biology.
3. Mnemons and the memorization of past signaling events. Reichert, P., and Caudron, F. (2021). Curr Opin Cell Biol 69, 127–135. PMID: 33618243
4. Prion-like proteins as epigenetic devices of stress adaptation. Oamen, H.P., Lau, Y., and Caudron, F. (2020). Experimental Cell Research 396, 112262. PMID: 32896568
5. Protein Phase Separation during Stress Adaptation and Cellular Memory. Lau, Y., Oamen, H.P., and Caudron, F. (2020). Cells 9, 1302. PMID: 32456195
6. Meeting report – shining light on septins. Caudron, F., and Yadav, S. (2018). Journal of Cell Science 131. PMID: 29360625
7. A Droplet to Sense Sugar Drops. Barral, Y., and Caudron, F. (2017). Molecular Cell 68, 1017–1019. PMID: 29272701
8. Spatial regulation of coalesced protein assemblies: Lessons from yeast to diseases. Saarikangas, J., and Caudron, F. (2017). Prion 259, 1–12. PMID: 28574744
9. Protein aggregation triggers a declining libido in elder yeasts that still have a lust for life. Caudron, F. (2017). Microbial Cell 4, 200–202. PMID: 28660204
10. Compartmentalization of ER-Bound Chaperone Confines Protein Deposit Formation to the Aging Yeast Cell. Saarikangas, J., Caudron, F., Prasad, R., Moreno, D.F., Bolognesi, A., Aldea, M., and Barral, Y. (2017). Current Biology 27, 773–783. PMID: 28262489
11. Aggregation of the Whi3 protein, not loss of heterochromatin, causes sterility in old yeast cells. Schlissel, G., Krzyzanowski, M.K., Caudron, F., Barral, Y., and Rine, J. (2017). Science 355, 1184–1187. PMID: 28302853
12. A role for the yeast CLIP170 ortholog, the plus-end-tracking protein Bik1, and the Rho1 GTPase in Snc1 trafficking. Boscheron, C., Caudron, F., Loeillet, S., Peloso, C., Mugnier, M., Kurzawa, L., Nicolas, A., Denarier, E., Aubry, L., and Andrieux, A. (2016). Journal of Cell Science 129, 3332–3341. PMID: 27466378
13. A sphingolipid-dependent diffusion barrier confines ER stress to the yeast mother cell. Clay, L., Caudron, F., Denoth-Lippuner, A., Boettcher, B., Buvelot Frei, S., Snapp, E.L., and Barral, Y. (2014). eLife 3, e01883. PMID: 24843009
14. [Sex and memory: what yeast can teach us]. Caudron, F., and Barral, Y. (2014). Médecine Sciences 30, 348–349. PMID: 24801024
15. Mnemons: encoding memory by protein super-assembly. Caudron, F., and , Barral, Y. (2014). Microbial Cell 1, 100–102. PMID: 28357228
16. A super-assembly of Whi3 encodes memory of deceptive encounters by single cells during yeast courtship. Caudron, F., and Barral, Y. (2013). Cell 155, 1244–1257. PMID: 24315096
17. Bud building by septin patch hole punching. Caudron, F., and Barral, Y. (2013). Developmental Cell 26, 115–116. PMID: 23906060
18. Mutation of Ser172 in yeast β tubulin induces defects in microtubule dynamics and cell division. Caudron, F., Denarier, E., Thibout-Quintana, J.-C., Brocard, J., Andrieux, A., and Fourest-Lieuvin, A. (2010). PloS One 5, e13553. PMID: 21042413
19. Oscillations in Cdc14 release and sequestration reveal a circuit underlying mitotic exit. Manzoni, R., Montani, F., Visintin, C., Caudron, F., Ciliberto, A., and Visintin, R. (2010). The Journal of Cell Biology 190, 209–222. PMID: 20660629
20. Septins and the lateral compartmentalization of eukaryotic membranes. Caudron, F., and Barral, Y. (2009). Developmental Cell 16, 493–506. PMID: 19386259
21. [A diffusion barrier allows budding yeast rejuvenation]. Caudron, F. (2009). Médecine Sciences 25, 217–218. PMID: 19361377
22. A new role for kinesin-directed transport of Bik1p (CLIP-170) in Saccharomyces cerevisiae. Caudron, F., Andrieux, A., Job, D., and Boscheron, C. (2008). Journal of cell science 121, 1506–1513. PMID: 18411245