

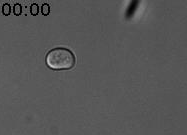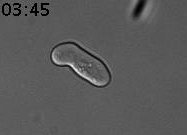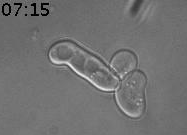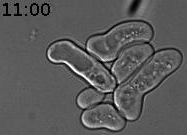
Figure 1. Cells were exposed to alpha-factor (7nM). The mother cell is at first shmooing and then only produces daughter cells, which shmoo. Time is in hours and minutes.
The pheromone refractory state depends on the inactivation of Whi3. Whi3 inhibits translation of mRNA encoding Cln3, the G1 cyclin required for the G1 to S phase transition. Whi3 contains prion-like domains that promote an inactivating conformational change upon pheromone treatment. We termed this type of protein mnemon. A mnemon is a protein that change of fold and super-assembly can encode a cellular memory. It is specifically induced by a stimulus and inherited asymmetrically during cell division (Figure 2).
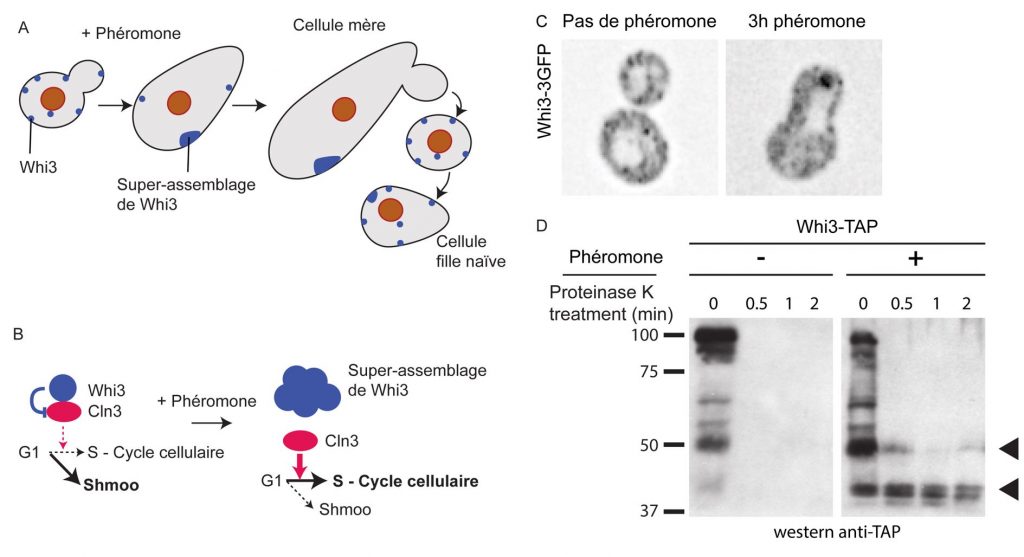
Figure 2. Schematic of the deceptive mating memory in budding yeast. A. A budding yeast cell exposed to pheromone shmoos. Whi3 super-assembles slowly, allowing the cell to escape the pheromone induced cell cycle arrest and produce a bud. The bud is born naive, free of Whi3 super-assembly and respond to pheromone. B. Upon pheromone treatment, Whi3 is inhibited through its super-assembly. The translation of Cln3 mRNA is no more repressed by Whi3, and Cln3 can commit cells to the S-phase. C. Whi3-3GFP localization in untreated and pheromone-exposed cells. Note the super-assembly of Whi3 after pheromone treatment. D. Biochemical characterization of Whi3’s conformation change. Whi3 becomes partially resistant to proteinase K treatment after cells were exposed to pheromone.
In contrast, prions are symmetrically inherited during cell division. We have identified that Whi3 association with the membranes of the endoplasmic reticulum (ER) promotes its retention in the mother cell. This is due to the compartmentalization of the ER membranes by lateral diffusion membranes at the bud neck, the limit between the mother cell and the future daughter cell. Disruption of the diffusion barrier leads to the transformation of Whi3 from a mnemon to a symmetrically inherited prion (Lau et al., Current Biology, 2022, Figure 3). Proteins with prion-like domains are very common in most living organisms and many of these molecules cause proteopathies such as neurodegenerative diseases or Creutzfeldt-Jakob disease. We test the idea that many proteins behave as mnemons, and we aim at understanding their biology in physiological and pathological contexts.
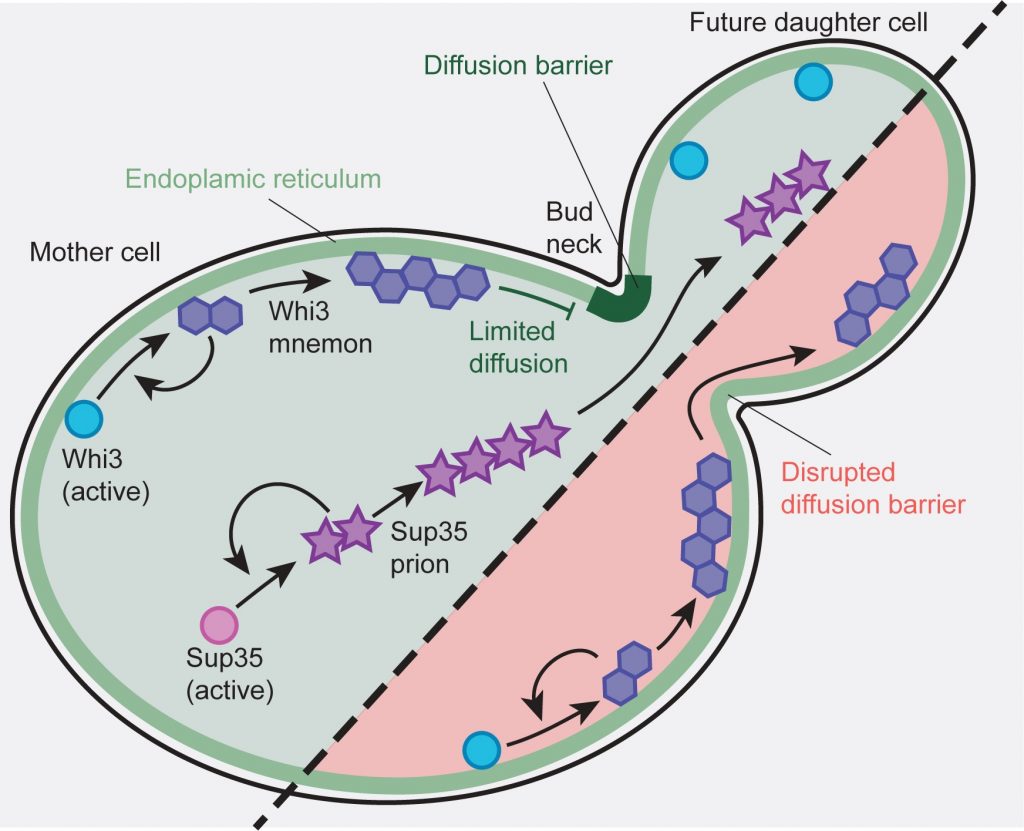
Figure 3. The Whi3 mnemon associates with endoplasmic reticulum membranes. A diffusion barrier at the bud neck restricts Whi3’s propagation to daughter cells. The prion form of Sup35, not associated with membranes is free to diffuse to daughter cells. When the diffusion barrier is genetically weakened, the Whi3 mnemon can propagate to daughter cells as a prion.

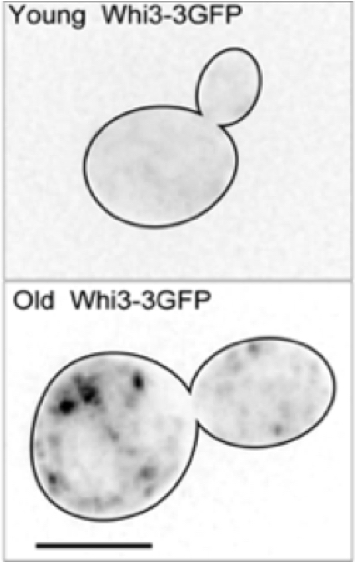
Figure 4. The left panel shows a movie of an old yeast cell expressing Hsp104-GFP. The age-induced protein deposit is seen as a dot that remains in the mother cell. The right panel shows a young yeast cell (top) expressing the mnemon Whi3-GFP. The signal is diffuse, while in an old yeast cell (bottom) it forms aggregates.
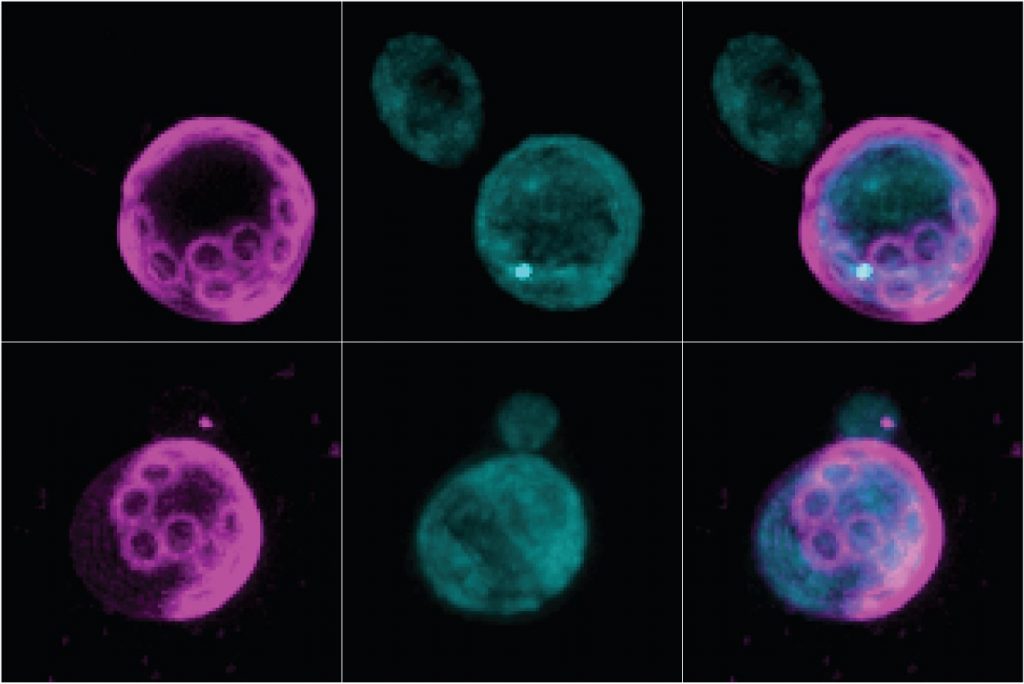
Figure 5. Bud scars (the rings, left) show the age of old cells. The protein Hsp104-GFP forms an age-induced aggregate (dot, middle upper panel) in untreated cells. Cells treated with tripentadecanoin (bottom) do not display this aggregate and live longer.







