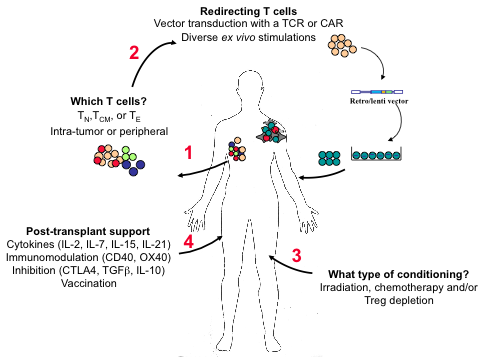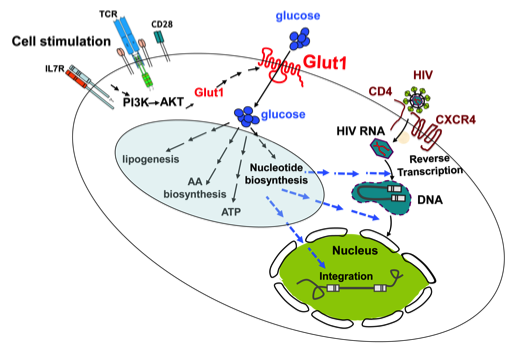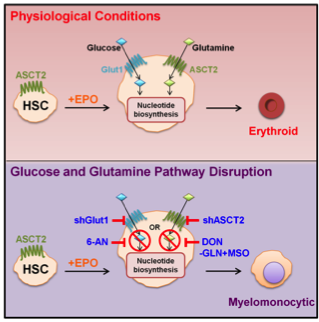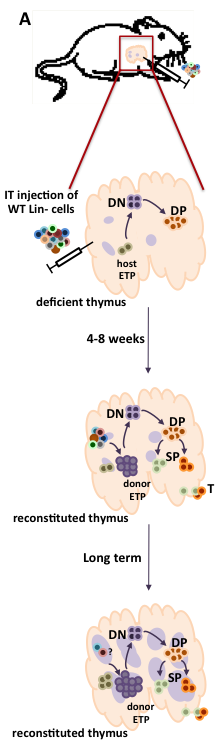
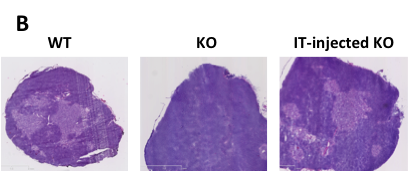
Figure 1: Fate of intrathymic progenitor cell transplants in immunodeficient mice with available thymic niches.
(A) WT hematopoietic progenitors, comprising lineage-negative (Lin-) cells, are intrathymically injected into immunodeficient mice with available thymic niches, such as that caused by ZAP-70-deficiency. These mice have only a rudimentary medulla (purple circles) and early thymocyte progenitors (ETP) are markedly reduced. Following intrathymic progenitor cell injection, thymopoiesis is restored by 4-8 weeks with increasing numbers of donor ETP as well as double negative (DN) immature progenitors, more mature CD4+CD8+ double positive (DP) thymocytes, as well as lineage committed CD4 and CD8 single positive (SP) thymocytes which emigrate from the thymus into the periphery. At long-term time points, the numbers of donor ETP remain elevated, fostering the continued differentiation of DP and SP thymocytes. (B) The direct intrathymic injection of hematopoietic progenitors into the thymus is associated with a long-term restoration of the thymic archicture. As shown in this hematoxylin-eosin staining of thymus sections from a WT, ZAP-70-KO (KO) and an IT-reconstituted ZAP-70-KO (IT-injected KO), intrathymic progenitor cell administration is associated with the presence of densely packed cortical regions and less dense medullary regions whereas KO mouse show defective medullary formation.